

Страница 2
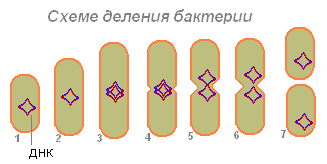
Основной способ размножения бактерий - деление клетки надвое (бинарное деление). При этом плазматическая мембрана и стенка впячиваются и перешнуровывают ее пополам. Впячивание мембраны происходит между точками прикрепления двух дочерних кольцевых молекул ДНК, в результате чего дочерние клетки обеспечиваются копиями материнской хромосомы. Бактерии обладают способностью к образованию эндоспор. Некоторые эндоспоры имеют плотные многослойные оболочки, устойчивы по отношению к агрессивным факторам внешней среды и длительно сохраняют способность к прорастанию.
Половой процесс у бактерий заключается в переносе ДНК от одной клетки к другой с последующей генетической рекомбинацией. Обмен наследственным материалом может происходить путем конъюгации (прямой контакт клеток), трансдукции (перенос ДНК вирусом-бактериофагом) или трансформации (поглощение фрагментов ДНК извне). Однако универсальным источником изменчивости являются мутации. В сочетании с темпом размножения бактерий они обеспечивают этим организмам высокую способность к адаптации к условиям внешней среды.
Различные виды бактерий могут использовать в качестве источника энергии почти любые органические соединения - не только питательные вещества, как сахара, аминокислоты и жиры, но и продукты выделения, например мочевину и мочевую кислоту, содержащиеся в моче, и вещества, входящие в состав экскрементов. Один из видов бактерий может использовать в качестве питательного субстрата даже пенициллин, убивающий многие бактерии.
Смотрите также
Основные факторы водной среды и их влияние на организмы
Введение
На нашей планете живые организмы освоили четыре среды обитания.
Водная среда была первой, в которой возникла и распространилась жизнь. Только
потом организмы овладели наземно...
Генная инженерия
Введение
Генная инженерия - это область биотехнологий, включающая в себя действия
по перестройке генотипов. Суть генной инженерии сводится к пониманию того, что
любой организм, будь т...
Асептика в биотехнологии
Введение
Биотехнологические процессы в основном проводят в асептических
условиях. Асептика - это комплекс мероприятий, направленных на предотвращение
попадания в среду посторонних вещ...
Некоторые микроорганизмы размножаются спорообразовани-ем (актиномицеты и грибы) и почкованием (дрожжи), у некоторых микроорганизмов наблюдается половое размножение, но большин-ство из них размножается бесполым (вегетативным) путем. При благоприятных условиях размножение протекает с необыкновен-ной быстротой — каждые 20-30 минут материнская бактериальная клетка делится на две дочерние. Дочерняя клетка со временем становится материнской и также делится. Таким образом, деление бактерий идет в геометрической прогрессии. Если бы такое деле-ние шло беспрепятственно, то через 48 часов одна бактерия могла бы дать потомство в сотни биллионов клеток, а через пять дней такую массу, которая заполнила бы собой бассейны всех морей и океанов. Однако этого не происходит, поскольку на микроорганиз-мы действуют различные факторы окружающей среды.
Делению клетки предшествует равномерное увеличение об-щего азота, РНК и белка в цитоплазме. Затем происходит реплика-ция (удвоение) ДНК. В делящейся клетке между спиралями ДНК разрываются водородные связи и образуются одиночные дочер-ние спирали ДНК (рис.25).
Рис. 25. Процесс бинарного деления па-лочковидных прокариот
3 -вытягивание клетки;
— формирование перегородки;
5 -разделение клеток.
Сразу после репликации ДНК начинается вытяжение клетки и образование поперечной перегородки за счет двух слоев цитоплазматической мембраны, выпячивающейся навстречу друг другу. Чаще всего перегородка образуется посередине материнской клетки, в результате чего дочерние клетки имеют примерно одинако-вые размеры. Между слоями перегородки идет формирование кле-точной стенки.
В процессе размножения одна из половин клетки постоянно сохраняет жгутики. На конечном этапе размножения бактерий жгу-тики вырастают и у другой половины.
Рост и размножение микроорганизмов зависит от различных факторов окружающей среды и видовых характеристик. Наблюде-ние за развитием микроорганизмов, культивируемых в жидкой пи-тательной среде в замкнутых резервуарах, показывает, что для роста биомассы необходимы наличие источника энергии, присут-ствие компонентов, необходимых для синтеза биомассы, отсутствие в среде ингибиторов, подавляющих рост клеток, поддержание в среде необходимых физико-химических условий. В этих условиях рост микроорганизмов условно можно подразделить на несколько последовательных фаз или периодов (рис. 26):

1. лаг-фаза (англ. lag — запаздывание) — период между посевом бактерий и началом размножения. В этот период происходит адап-тация бактериальной культуры к питательной среде. Она проявля-ется в накоплении оптимального количества необходимых фермен-тов, в инактивации некоторого ингибитора, присутствующего в сре-де, в прорастании спор и др. При благоприятных условиях бактерии увеличиваются в размерах и готовятся к делению. Лаг-фаза мо-жет длиться от 10 минут до нескольких часов, но в среднем она составляет 4-5 часов.
3. Фаза логарифмического или экспоненциального роста явля-ется периодом наиболее интенсивного деления бактерий. Бактерии делятся каждые 20-40 минут. Во время этой фазы бактерии осо-бенно ранимы, что объясняется высокой чувствительностью рас-тущих клеток к факторам окружающей среды. Продолжительность экспоненциального роста зависит от концентрации питательных ве-ществ в субстрате и в среднем составляет 5-6 часов.
5. Фаза стационарного роста вызывается постепенным исто-щением среды, накоплением в ней литических ферментов, хими-ческим ингибированием роста микробной клетки продуктами ме-таболизма. Эта фаза отличается от предыдущей повышенной со-противляемостью бактерий многим химическим и физическим фак-торам. К началу этой фазы количество жизнеспособных клеток достигает максимального уровня и остается на этом максимуме в течение нескольких часов в зависимости от вида микроорганизмов и особенностей их культивирования. В конце этой фазы у некоторых микроорганизмов наблюдается процесс спорообразования.
6. Завершающая фаза процесса размножения — фаза старения и гибели — характеризуется отмиранием бактерий из-за истощения питательной среды и накопления в ней продуктов метаболизма. Наблюдается автолиз микроорганизмов как экстремальное прояв-ление нестабильности клетки после прекращения роста. Продол-жительность этой фазы может составлять от нескольких часов до нескольких недель.
Дата публикования: 2015-11-01; Прочитано: 2315 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.002 с)…
Бактерии, как и все живые организмы, размножаются. Проис-ходит это чаще всего путем простого поперечного деления в раз-личных плоскостях. При этом образуются разнообразные сочетания клеток: парные соединения, одиночные клетки, гроздья, цепочки, пакеты и др.
Некоторые микроорганизмы размножаются спорообразовани-ем (актиномицеты и грибы) и почкованием (дрожжи), у некоторых микроорганизмов наблюдается половое размножение, но большин-ство из них размножается бесполым (вегетативным) путем.
При благоприятных условиях размножение протекает с необыкновен-ной быстротой — каждые 20-30 минут материнская бактериальная клетка делится на две дочерние. Дочерняя клетка со временем становится материнской и также делится.
Таким образом, деление бактерий идет в геометрической прогрессии. Если бы такое деле-ние шло беспрепятственно, то через 48 часов одна бактерия могла бы дать потомство в сотни биллионов клеток, а через пять дней такую массу, которая заполнила бы собой бассейны всех морей и океанов. Однако этого не происходит, поскольку на микроорганиз-мы действуют различные факторы окружающей среды.
Делению клетки предшествует равномерное увеличение об-щего азота, РНК и белка в цитоплазме.
Затем происходит реплика-ция (удвоение) ДНК. В делящейся клетке между спиралями ДНК разрываются водородные связи и образуются одиночные дочер-ние спирали ДНК (рис.25).
25. Процесс бинарного деления па-лочковидных прокариот
1 — образование одиночных спиралей ДНК;
2 — удвоение (репликация) ДНК;
3 -вытягивание клетки;
— формирование перегородки;
4 — окончание формирования перего-родки и образование выпуклой клеточной стенки;
5 -разделение клеток.
Сразу после репликации ДНК начинается вытяжение клетки и образование поперечной перегородки за счет двух слоев цитоплазматической мембраны, выпячивающейся навстречу друг другу.
Чаще всего перегородка образуется посередине материнской клетки, в результате чего дочерние клетки имеют примерно одинако-вые размеры. Между слоями перегородки идет формирование кле-точной стенки.
Одиночная спираль ДНК в новых клетках служит матрицей для создания второй спирали, в результате чего образуется двой-ная спираль ДНК с восстановленными водородными связями и формируется новый нуклеоид.
В процессе размножения одна из половин клетки постоянно сохраняет жгутики.
На конечном этапе размножения бактерий жгу-тики вырастают и у другой половины.
Рост и размножение микроорганизмов зависит от различных факторов окружающей среды и видовых характеристик. Наблюде-ние за развитием микроорганизмов, культивируемых в жидкой пи-тательной среде в замкнутых резервуарах, показывает, что для роста биомассы необходимы наличие источника энергии, присут-ствие компонентов, необходимых для синтеза биомассы, отсутствие в среде ингибиторов, подавляющих рост клеток, поддержание в среде необходимых физико-химических условий.
В этих условиях рост микроорганизмов условно можно подразделить на несколько последовательных фаз или периодов (рис. 26):
Рис. 26. Типичная кривая роста популяции микроорганизмов 1 — лаг-фаза;
2 — фаза ускоренного роста; 3 — фаза логарифмического (экспоненциального) роста;
4 — фаза замедления роста; 5 — фаза стационар-ного роста; 6 — фаза старения и отмирания.
лаг-фаза (англ. lag — запаздывание) — период между посевом бактерий и началом размножения. В этот период происходит адап-тация бактериальной культуры к питательной среде. Она проявля-ется в накоплении оптимального количества необходимых фермен-тов, в инактивации некоторого ингибитора, присутствующего в сре-де, в прорастании спор и др. При благоприятных условиях бактерии увеличиваются в размерах и готовятся к делению.
Лаг-фаза мо-жет длиться от 10 минут до нескольких часов, но в среднем она составляет 4-5 часов.
2. Фаза ускоренного роста наблюдается после лаг-фазы и характеризуется нарастанием темпов деления микроорганизмов и накопления биомассы.
3. Фаза логарифмического или экспоненциального роста явля-ется периодом наиболее интенсивного деления бактерий.
Бактерии делятся каждые 20-40 минут. Во время этой фазы бактерии осо-бенно ранимы, что объясняется высокой чувствительностью рас-тущих клеток к факторам окружающей среды. Продолжительность экспоненциального роста зависит от концентрации питательных ве-ществ в субстрате и в среднем составляет 5-6 часов.
4. Фаза замедления роста является переходным периодом от экспоненциального роста к фазе стационарного роста. Во время этой фазы наблюдается истощение питательных веществ субстрата и накопление в нем продуктов метаболизма, что снижает интенсив-ность размножения микроорганизмов.
Фаза стационарного роста вызывается постепенным исто-щением среды, накоплением в ней литических ферментов, хими-ческим ингибированием роста микробной клетки продуктами ме-таболизма. Эта фаза отличается от предыдущей повышенной со-противляемостью бактерий многим химическим и физическим фак-торам. К началу этой фазы количество жизнеспособных клеток достигает максимального уровня и остается на этом максимуме в течение нескольких часов в зависимости от вида микроорганизмов и особенностей их культивирования.
В конце этой фазы у некоторых микроорганизмов наблюдается процесс спорообразования.
6. Завершающая фаза процесса размножения — фаза старения и гибели — характеризуется отмиранием бактерий из-за истощения питательной среды и накопления в ней продуктов метаболизма. Наблюдается автолиз микроорганизмов как экстремальное прояв-ление нестабильности клетки после прекращения роста.
Продол-жительность этой фазы может составлять от нескольких часов до нескольких недель.
Дата публикования: 2015-11-01; Прочитано: 2316 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.001 с)…
Размножение микроорганизмов - бинарное деление одноклеточных микроорганизмов (бактерий, риккетсий, простейших, дрожжей), в результате которого образуются две новые дочерние полноценные особи, наделенные генетической информацией материнской клетки. Дрожжеподобные грибы могут размножаться почкованием, спорами; плесневые грибы и актиномицеты размножаются обычно спорами.
Бактерии
Размножаются простым поперечным делением.
Бактерии являются гаплоидными клетками. В состав бактериальной клетки входит капсула, клеточная стенка, цитоплазматическая мембрана, цитоплазма, где располагаются мезосомы, рибосомы, нуклеоид, и включения. Некоторые бактериальные клетки имеют жгутики и образуют споры.
В отличие от животных клеток такие внутренние структуры бактериальной клетки, как мезосомы, рибосомы, нуклеоид, не имеют мембран, отграничивающих их от цитоплазмы.
По способу питания бактерий делят на автотрофов и гетеротрофов, по способу дыхания - на аэробов и анаэробов.
Актиномицеты
Размножаются спорами и поперечным делением (отшнуровыванием) гиф.
Занимают промежуточное положение между грибами и бактериями. Среди лучистых грибов сеть патогенные виды - возбудители актиномикозов. Многие актиномицеты являются продуцентами антибиотиков. (см.
Антибиотики). В «Определителе» Берджи актиномицеты названы стрептомицетами.
Дрожжи
Существует 2 вида размножения дрожжей - вегетативное (бесполое) и половое с образованием спор. У большинства видов дрожжей вегетативное размножение осуществляется почкованием, редко делением (Schizosaccharomyces). Аспорогенные. дрожжи размножаются только почкованием. Половое размножение происходит при неблагоприятных условиях, когда дрожжи перестают почковаться и превращаются в сумки (аски) со спорами - аскоспоры.
Половой процесс заключается в копуляции (слиянии) 2 вегетативных клеток путем сближения их и образования копуляционного канала, в котором происходит слияние частей плазмы и ядра клеток, называемое кариогамией, с образованием диплоидной зиготы, представляющей 2 клетки, соединенные копуляционным каналом.
Редукционное деление, или мейоз, сопровождаемое уменьшением числа хромосом вдвое, происходит сразу, без полового процесса, и зигота превращается в аск с 4 гаплоидными спорами, поэтому вегетативное поколение таких спор гаплоидно. Споры прорастают без копуляции. Так происходит размножение у дрожжей Zygosaccharomyces. У дрожжей Saccharomyces половой процесс происходит при слиянии спор или проросших из них клеток с образованием диплоидной зиготы, которая сразу начинает почковаться, образуя диплоидное потомство.
Мейоз происходит непосредственно перед образованием спор.
Плесневые грибы
У Грибов различают вегетативное, половое и бесполое размножение.
Вегетативное размножение может осуществляться при отделении от основной массы мицелия его частей, которые могут развиваться самостоятельно, а так же путем почкования мицелия или отдельных клеток у дрожжевых грибов.
Половое размножение состоит в слиянии половых клеток, в результате чего возникает зигота.
Бесполое размножение осуществляется при помощи специальных образований, называемых спорами. Споры могут развиваться внутри специальных споровместилищ или на концах особых выростов мицелия – конидиеносцах.
Основной способ размножения плесневых грибов – при помощи спор. Плесень размножается невероятно быстро.
В обыкновенной хлебной плесени можно различить маленькие чёрные точки — спорангии, в которых образуются споры. В одном спорангии содержится до 50.000 спор, каждая из которых способна воспроизвести сотни миллионов новых спор всего за несколько дней! А если условия благоприятные, плесень быстро появится на книге, обуви или на упавшем дереве в лесу.
Бактерии: Жизнедеятельность бактерий характеризуется ростом - фор-мированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размноже-нием - самовоспроизведением, приводящим к увеличению ко-личества бактериальных клеток в популяции.
Бактерии размножаются путем бинарного деления пополам, реже путем почкования.
Актиномицеты, как и грибы, могут размножаться спорами. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Актиномицеты, являясь ветвящимися бактериями, размножаются путем фрагментации нитевидных клеток Грамположительные бактерии делятся путем врастания синтези-рующихся перегородок деления внутрь клетки, синтезируют поперечную перегородку от периферии к центру при участии мезосом.
а грамотрицательные - путем перетяжки(на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь.), в результате образования гантелевидных фигур, из которых образуются две одинаковые клетки. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних.
У других бактерий кроме размножения наблюдается половой процесс, но в самой примитивной форме.
Половой процесс бактерий отличается от полового процесса эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового процесса, а именно обмен генетическим материалом, происходит и в этом случае. Это называется генетической рекомбинацией.
Делению клеток предшествует репликация бактериальной хро-мосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной ни-тью), приводящая к удвоению молекул ДНК бактериального ядра - нуклеоида. Репликация ДНК происходит в три этапа: инициация, элон-гация, или рост цепи, и терминация.
Размножение спирохет: поперечное деление-деление клетки у бактерий, при котором материнская клетка дает начало двум дочерним клеткам. Осуществляется в три стадии:
1) реплика-ция молекулы ДНК кольцевой хромосомы, присоединенной к мезосоме, которая так же делится на две части;
2) разведение с помощью мезосом двух дочерних кольцевых хромосом;
3) раз-деление цитоплазмы поперечной перегородкой, которая образу-ется от периферии к центру клетки.
Размножение грибов:
Большинство грибов способно к вегетативному, собственно бесполому и половому размножению.
Характерен плеоморфизм - наличие одновременно нескольких видов спороношений, например, бесполого и полового.
Вегетативное размножение
- Частями мицелия.
- Специализированными образованиями: артроспорами (оидиями) с тонкими стенками или хламидиоспорами с толстыми, образуются они, с некоторыми отличиями, при распаде мицелия на части, а затем дают начало новому.
- Почкование гиф или отдельных клеток (например, у дрожжей).
Также почкуются аскоспоры у сумчатых и базидиоспоры у головнёвых. Образующиеся почки постепенно отделяются, растут и со временем сами начинают почковаться.
Бесполое размножение
Собственно бесполое размножение идёт посредством спор.
В зависимости от способа образования различают эндогенные и экзогенные споры.
- Эндогенные споры (спорангиоспоры) характерны для низших грибов.
Образуются внутри особых клеток, называемыхспорангиями.
- Экзогенные споры обычно называют конидиями,они имеются у высших и у некоторых низших грибов.
Образуются на вершинах или сбоку специальных гиф - конидиеносцев, ориентированных вертикально, которые могут быть простыми или разветвлёнными.
Покрыты плотной оболочкой, поэтому довольно устойчивы, но неподвижны. Могут подхватываться воздушными потоками или животными и переноситься на значительные расстояния. При прорастании дают ростовую трубку, а затем гифы.
Половое размножение

Конъюгация гамет
Для низших грибов свойственно слияние гаплоидных гамет путём изогамии, анизогамии (гетерогамии) или оогамии.
В случае оогамии развиваются половые органы - оогонии (женские) и антеридии (мужские). При оплодотворении происходит образование ооспоры - это зигота, которая покрывается толстой оболочкой, некоторое время проводит в состоянии покоя, после чего прорастает.
Скорость и фазы размножения бактерий в стационарных условиях.
При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост культуры.
Рост периодической культуры бактерий, выращиваемых на жидкой питательной среде, подразделяют на несколько фаз, или периодов:
1. лаг-фаза;
2. фаза логарифмического роста;
3. фаза стационарного роста, или максимальной концентрации бактерий;
4. фаза гибели бактерий.
Эти фазы можно изобразить графически в виде отрезков кри-вой размножения бактерий, отражающей зависимость логариф-ма числа живых клеток от времени их культивирования.
Лаг-фаза - период между по-севом бактерий и началом размножения.
Продолжительность лаг-фазы в среднем 4-5 ч. Бактерии при этом увеличиваются в раз-мерах и готовятся к делению; нарастает количество нуклеино-вых кислот, белка и других компонентов.
Фаза логарифмического (экспоненциального) роста является периодом ин-тенсивного деления бактерий. Продолжительность ее около 5- 6 ч. При оптимальных условиях роста бактерии могут делиться каждые 20-40 мин.
Во время этой фазы бактерии наиболее ра-нимы, что объясняется высокой чувствительностью компонен-тов метаболизма интенсивно растущей клетки к ингибиторам синтеза белка, нуклеиновых кислот и др.
Затем наступает фаза стационарного роста, при которой количество жиз-неспособных клеток остается без изменений, составляя макси-мальный уровень (М-концентрация). Ее продолжительность вы-ражается в часах и колеблется в зависимости от вида бактерий, их особенностей и культивирования.
Завершает процесс роста бактерий фаза гибели, характеризующаяся отмиранием бак-терий в условиях истощения источников питательной среды и накопления в ней продуктов метаболизма бактерий. Продолжи-тельность ее колеблется от 10 ч до нескольких недель. Интен-сивность роста и размножения бактерий зависит от многих фак-торов, в том числе оптимального состава питательной среды, окислительно-восстановительного потенциала, рН, температуры и др.
Скорость роста бактерий зависит как от внешних условий, так и от физиологических особенностей самой клетки.
При наличии благоприятных условий рост бактериальной клетки завершается размножением. Основным способом размножения большинства бактерий является простое деление клетки пополам. Делению предшествует репликация (удвоение) хромосомы. Эти два процесса тесно взаимосвязаны. Частота репликации регулируется скоростью роста клетки. Репликация бактериальной хромосомы осуществляется описанным ранее способом (см. п. 3.2.5).
Изучение закономерности равномерного распределения генетического материала между дочерними клетками, образовавшимися в результате деления материнской клетки, позволило Г. Жакобу, С. Бреннеру и Т. Кузену (1963) сформулировать концепцию репликона. Репликон — единица репликации, это участок ДНК, содержащий регуляторные элементы, необходимые для независимой репликации. У бактерий таковым являются хромосома и плазмиды. Каждый репликон содержит не менее двух локусов, участвующих в контроле репликации: структурный ген-репликатор (ген-инициатор), детерминирующий синтез белка-инициатора и специальный сайт-репликатор, который распознает сигналы на начало удвоения хромосомы.
После некоторого периода роста клетка достигает определенного физиологического состояния. Из цитоплазматической мембраны в репликон поступают сигналы о необходимости репликации хромосомы и готовности клетки к делению. Под влиянием сигналов активизируется деятельность структурного гена и синтезируется белок-инициатор.
Он, воздействуя на репликатор, запускает репликацию.
Между системой репликации хромосомы и делением клетки существует координированное взаимодействие: делению клетки всегда предшествует удвоение хромосомы. После завершения репликации начинается процесс деления клетки. У грамположительных бактерий и цианобактерий это осуществляется образованием поперечной перегородки, разделяющей материнскую клетку на две равноценные дочерние.
Деление происходит следующим образом.
Вначале
синтезируется двуслойная цитоплазматическая мембрана. Затем на внутренней стороне клеточной стенки образуются два бугорка. Они интенсивно растут и, проникая кольцеобразно внутрь клетки между слоями образовавшейся цитоплазматической мембраны, образуют двойную перегородку, делящую клетку пополам.
Деление большинства грамотр тщательных бактерий
происходит путем перетяжки. При этом геномы расходятся по полюсам клетки, цитоплазматическая мембрана и клеточная стенка растягиваются, впячиваясь от периферии к центру клетки до контакта друг с другом. В результате клетка перешнуровывается на две дочерние. Деление клеток образованием перегородки или перетяжкой получило название бинарного в связи с формированием двух одинаковых дочерних клеток.
Кроме описанного бинарного деления, у бактерий известен другой способ размножения * почкование. Почкованием размножаются бактерии родов Hyphomicrobium, Pedomicrobium и других, объединенных в группу почкующихся бактерий.
Эти организмы имеют вид вытянутых палочек (0,5х 2 мкм), иногда грушевидных, оканчивающихся гифами, или простеками (выростами).
Размножение у этих бактерий начинается с образования почки на конце гифы или непосредственно на материнской клетке.
Почка разрастается в дочернюю клетку, формирует жгутик и отделяется от материнской клетки. По достижению зрелого состояния жгутик теряется и процесс развития повторяется.
В отличие от бинарного деления при почковании исходная клетка остается материнской, а вновь образованная — дочерней.
Между ними имеются морфологические и физиологические различия.
Актиномицеты размножаются фрагментами мицелия и спорами. У одних (род Micromonospora) единичные споры формируются на гифах вегетативного мицелия, у других (род Streptomyces и др.) цепочки спор образуются на концах гиф воздушного мицелия, так называемых конидиеносцах.
Фрагменты мицелия и споры в благоприятных условиях влажности, температуры прорастают и дают начало новым организмам.
Нитчатые цианобактерии кроме бинарного деления размножаются участками трихом и гормогониями. Последние представляют собой укороченные нити, состоящие из мелких вегетативных клеток одинаковой формы и размеров. При отмирании средних клеток трихома (нити) гормогонии выскальзывают из чехла материнского трихома, растут, делятся, образуя новые трихомы.
Гормогонии, в отличие от материнского трихома, не имеют гетероцист и никогда не окружены чехлом.
Независимо от того, каким путем идет процесс размножения бактерий, скорость этого процесса огромна: за 24 ч может смениться столько поколений, сколько у человека за пять тысяч лет.
Скорость размножения зависит от многих условий и для каждого вида бактерий различна. При наличии в среде необходимых питательных веществ, благоприятной температуры и кислотности среды деление каждой клетки может повторяться через 20-30 мин (Е. coli). При такой скорости размножения из одной клетки за сутки возможно образование 472 * 1019 клеток (273, 72 генерации).
Интенсивное размножение имеет для бактерий большое биологическое значение. Оно обеспечивает сохранение микроорганизмов на земной поверхности. При наступлении неблагоприятных условий они погибают массами, но достаточно сохраниться где-нибудь нескольким клеткам, как при подходящих условиях они дадут большое потомство клеток.
Численность популяции микроорганизмов в естественных местообитаниях, например, в почве или воде, постоянно меняется в соответствии с изменением условий существования.
Но в лабораторных условиях на питательных средах изменение численности популяции микроорганизмов происходит закономерным образом.
А так же в разделе «РАЗМНОЖЕНИЕ БАКТЕРИЙ»
Актиномицеты (Actinomyces) в переводе с латинского — лучистый гриб, обособленная группа микроорганизмов обладающая рядом морфологических признаков низшего вида гриба и не образующей спор бактерии.
Морфология актиномицетов
Строение актиномицитов имеет схожие черты с мицелиальными грибами, грифы мицелия имеют толщину в среднем 0.7 мк, изменяющуюся в пределах 0,5-1,2 мм, что на много меньше чем у грибов.
Для нитей, прямого или немного изогнутого вида, не имеющих поперечных перегородок, характерно моноподиальное или в отдельных случаях мутовчатое ветвление. Клеточная оболочка по составу имеет ряд черт грамположительных бактерий.
Размножение актиномицетов
Actinomyces размножаются с помощью субстрационного мицелия прорастающего в субстрате и воздушного мицелия вырастающего с плодоносцев-спорангиофоров.
Плодоносцы в зависимости от вида имеют различную форму завитков от закрученной формы, до прямой или волнистой.
Некоторые виды актиномицетов имеют спороносные ветки расположенные в виде мутовок или пучков, часто они моноподиально висят на нитях мицелия.
Образование спор происходит с помощью фрагментации или сегментации.
Фрагментация — это процесс дробления протопласта спороносной ветки на сто и более меленьких комков содержащих базофильное а также ядерное вещество.
Комки, превращаясь в споры, располагаются длинной цепочкой в спороносце.
Сегментация — это процесс разделения спороносца на сегменты с палочковидной формой, с помощью поперечных перегородок, они округляются и преобразовываются в споры.
Оболочки спор у разных видов имеют гладкую иногда бугристую, зубчатую, шиповидную, волосистую поверхность. Выросты на поверхности оболочек хорошо видны через электронный микроскоп.
В большинстве случаев актиномицеты аэрофилы и мезофилы, но получили распространение и термофилы, многие их виды способны образовывать пигменты разных цветов.
Актиномицеты, имея разнообразный набор ферментов способны синтезировать различные вещества и выделять их большими количествами в окружающую среду. Среди этих веществ, обладающих высокой физиологической активностью, встречаются многие витамины, некоторые аминокислоты, токсины, каротиноиды, фитогормоны и другие.
Также стоит упомянуть способность актиномицетитов образовывать разнообразные виды антибиотиков.
Одной из функций жизнедеятельности прокариотов, как и любых других живых существ, является размножение. По своей сути, процесс размножения бактерий можно охарактеризовать как рост числа особей, что происходит благодаря делению бактерий.
Современная микробиология описала схемы митоза, мейоза и амитоза – так делятся эукариоты, а прокариоты размножаются прямым делением.
Прокариоты размножаются преимущественно путем деления материнской бактериальной клетки на 2 идентичные дочерние. При благоприятных условиях бинарное деление происходит каждые 20 минут, а в случае ухудшения условий окружающей среды время, необходимое клетке для роста и деления, увеличивается. В случае неблагоприятных внешних условий прокариоты прекращают размножение на время или вовсе.
Непосредственно самому процессу разделения клетки пополам предшествует период роста цитоплазмы и репликации (удвоения) хромосомы бактерии, как на фото.

Репликации кольцевой бактериальной хромосомы
Увеличение клеточных размеров происходит вследствие целого ряда скоординированных процессов биосинтеза, которые жестко контролируются. Процесс роста бактерии не бесконечен – по достижению прокариотом заданных критических размеров происходит деление.
Механизм репликации бактериальной ДНК
При удвоении ДНК нуклеоида (аналог ядра в бактериальной клетке) реализуется следующая схема:
- инициация – начало деления ДНК под действием репликона (ферментативного аппарата, участка ДНК, содержащего информацию о дублировании);
- элонгация – удлинение, рост хромосомной цепи;
- терминация – завершение роста цепи и спирализация ДНК при репликации.
Параллельно с репликацией ДНК происходит рост самой клетки, и расстояние между прикрепленными посредством мезосом к цитоплазматической мембране двумя новыми хромосомами постепенно увеличивается. Прокариотическая клетка начинает делиться спустя некоторое время после репликации. Очевидно, именно дублирование ДНК запускает процесс разделения.
Подобный процесс отсутствует для мейоза эукариотов. Процесс мейоза во многом отличается от размножения прокариотов. Кроме того, разделение материнской клетки на две части для грамположительных и грамотрицательных бактерий имеет свои особенности.
Размножение грамотрицательных бактерий
Грамотрицательные бактерии имеют сравнительно тонкую клеточную стенку, на которой приблизительно по центру расположена кольцевая органелла – септальное кольцо. Разделение бактерий происходит путем сокращения органеллы и формирования перетяжки между дочерними клетками, что видно на фото.

Септальное кольцо представляет собой сложный белковый комплекс, куда входит более 12 различных белков. Оно формируется путем последовательного присоединения белков друг к другу в строгой последовательности.
Белки септального кольца выполняют следующие функции, необходимые для размножения:
- моделируют присоединение филаментов (белков кольца) в определенной последовательности к Z-кольцу (незрелая форма кольцевой органеллы);
- обеспечивают привязку Z-кольца к мембране;
- координируют формирование кольцевой органеллы с сегрегацией (разъединением) хромосомы;
- синтезируют пептидогликан – наиболее значимый компонент бактериальной клеточной стенки, который осуществляет осмотическую защиту;
- осуществляют гидролиз пептидогликана для разделения новых клеток.
Перетяжка у грамотрицательных бактерий охватывает все оболочки клетки – цитоплазматическую (внутреннюю) и внешнюю мембраны, а также связанный с ними липопротеидом тонкий слой пептидогликана.
Во время мейоза эукариотов подобный метод разделения клетки перетяжкой не встречается.
Размножение грамположительных бактерий
Толщина стенки грамположительной бактерии более чем в два раза превышает толщину стенки у бактерий грамотрицательных.
Процесс размножения грамположительной бактерии не имеет сходства с митозом и отличается от мейоза эукариотов. По окончании процесса репликации ДНК грамположительные бактерии не создают перетяжку, а синтезируют поперечную перегородку, как на фото. В процессе синтеза, как и у грамотрицательных бактерий при образовании перетяжки, принимают участие мезосомы, формируя перегородку от края к центру клеточной структуры.

Поперечное бинарное деление бактериальной прокариотической клетки всегда продольно и поперечно симметрично, что является еще одним отличием процесса от мейоза клеточной структуры эукариотов.
При благоприятных условиях прямое бинарное деление бактериальных клеток может осуществляться как одной, так и в нескольких плоскостях, что невозможно для мейоза. В случае, когда клетки после разделения не расходятся, происходит образование различных по форме объединений:
- при сечении клетки в одной плоскости формируются цепочки шаровидных или палочковых клеток (сферические диплококки, цепочка палочковидных бактерий, как на фото);
- при разделении в разных плоскостях наблюдаются клеточные скопления разнообразных форм (цепочки стрептококков, пакеты сарцин, грозди стафилококков).
Разнообразие форм прокариотов, которое видно на фото, совершенно не реализуемо для мейоза ядерных клеток.
Подобное поперечное разделение характерно не только для грамположительных бактерий, но и для нитчатых цианобактерий.
Множественное деление цианобактерий
Одной из разновидностей бинарного размножения прокариотов является множественное образование дочерних прокариотов из материнской клетки, типичное для цианобактерий, и совершенно не характерное для мейоза.

А — размножение цианобактерий рода Dermocarpa
Б — размножение цианобактерий рода Chroococcidiopsis
Первоначально происходит рост цитоплазмы и репликация хромосомы. Затем, как видно на видео, внутри дополнительного фибриллярного слоя материнского организма осуществляются последовательные бинарные деления, которые приводят к образованию баеоцитов (маленьких клеток). Их число может колебаться от 4 до 1000 единиц и связано с видом цианобактерии. Высвобождаются баеоциты после разрыва стенки материнского прокариота, что видно на видео.
Помимо равновеликого разделения, некоторые бактерии размножаются почкованием.
Почкование как частный случай бинарного деления
У фото- и хемотрофов, независимо от источника пищи (автотрофы или гетеротрофы), обнаруживается возможность размножения организма почкованием.
Механизм процесса выглядит следующим образом:
- на полюсе материнской клетки формируется почка;
- происходит рост почки до размеров материнского организма (это видно на фото), причем для почки синтезируется новая клеточная стенка;
- полноценная дочерняя клетка отделяется от материнской.
Если процесс бинарного разделения не имеет ограничений, как в случае мейоза
для эукариотов, то почкование зависит от факта старения прокариота. В среднем материнская клетка отделяет не более 4 почек.
Почкование имеет свои специфические особенности:
- сохраняется только продольная симметрия (хорошо видно на фото);
- после почкования получаются материнская и дочерняя клетки, тогда как после бинарного разделения материнской клетки нет – есть две равноценные дочерние;
- материнский и дочерний организмы не идентичны, различия между ними хорошо видны – наблюдается процесс старения.
При благоприятных физико-химических условиях прокариоты способны делиться в геометрической прогрессии и заполонить собой весь мир. Однако на деле такого не происходит, так как существуют факторы, сдерживающие бактериальное деление.
Факторы, ограничивающие деление
При всем видовом разнообразии и приспособленности бактерии не размножаются бесконечно. Исследования показал, что рост популяции бактерий происходит в согласии с законом размножения микроорганизмов и поддается числовому и графическому описанию.
Рост популяции, связанный с делением бактерий, состоит из нескольких фаз:
- лаг-фаза – период адаптации, когда требуется время для приспособления к новым условиям обитания, деление не имеет высокого значения;
- логарифмическая фаза – период с наибольшим количеством делений и экспоненциальным ростом популяции;
- стационарная фаза – время, когда рост колонии бактерий стремится к нулю, деление бактерий уравнивается с количеством погибших из-за ограниченности пищевых ресурсов;
- замедление роста – происходит из-за значительного сокращения пищевых ресурсов и накопления токсичных продуктов жизнедеятельности.
Неблагоприятные условия провоцируют прекращение деления бактерий и, как следствие, неизбежную гибель популяции.

Работаю врачом ветеринарной медицины. Увлекаюсь бальными танцами, спортом и йогой. В приоритет ставлю личностное развитие и освоение духовных практик. Любимые темы: ветеринария, биология, строительство, ремонт, путешествия. Табу: юриспруденция, политика, IT-технологии и компьютерные игры.
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.
После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
На заключительной стадии деления клеточная оболочка сжимается и разрушается или участвует в синтезе септы с последующим автолизом, образуя две отдельных клетки
Процесс деления у бактерий контролируется гомологом тубулина, белком FtsZ, который образует кольцевую структуру в месте деления
Вместе с FtsZ, в месте деления образуется набор, состоящий примерно из 8 белков, участвующих в делении
Место деления клетки определяется двумя системами отрицательной регуляции: блокирующим эффектом нуклеоида и системой Min
Большинство клеток эукариот делятся точно посередине, образуя две одинаковые дочерние клетки. Деление скоординировано с завершением репликации и сегрегацией хромосом. Обычно деление происходит по завершению периода роста, во время которого масса клеток удваивается. После сегрегации хромосом наступает цитокинез, в результате которого клетка разделяется на две. Во время цитокинеза все слои клеточной мембраны локально принимают кольцеобразную форму. Как показано на рисунке ниже, цитокинез осуществляется, по крайней мере, двумя различными путями.
У грамотрицательных микроорганизмов , таких как Е. coli, деление происходит при сокращении слоев существующей оболочки, с последующим разрывом образующейся перемычки. У других бактерий, например у грамположительных В. subtilis, новообразованные кольцевые структуры материала клеточной стенки растут внутрь клетки, образуя перегородку. Когда образование перегородки завершилось, между сестринскими клетками образуется двойная мембрана, но клетки остаются связанными друг с другом. Разделение клеток представляет собой самостоятельное событие, которое включает в себя автолиз материала перегородки. В зависимости от условий роста, автолиз перегородки может происходить достаточно медленно и сопровождаться возникновением длинных цепей связанных между собой клеток.
При выделении и характеристике мутантов fis (филаментарные температурочувствительные мутации) был идентифицирован ряд генов, необходимых для деления. Клетки мутантов fis при непермиссивной температуре растут в виде длинных неделящихся филаментов. У большинства бактерий обнаружено около 8 генов fis. Плодотворным оказалось наблюдение Люткенхауза, который обнаружил, что белок FtsZ образует кольцеобразные структуры непосредственно под клеточной мембраной на месте деления. Затем к этому «Z-кольцу» в определенном порядке подходят остальные белки деления. Этот процесс для клеток Е. coli представлен на рисунке ниже. Функции большинства этих белков неизвестны.
Ключевой белок деления, FtsZ , представляет собой гомолог тубулина эукариот, белка, входящего в состав цитоскелета и формирующего микротрубочки. Подобно тубулину, этот белок является ГТФазой и в присутствии ГТФ полимеризуется с образованием линейных прото-филаментов, in vitro формирующих пучки и плоские структуры. Кольцевая структура белка FtsZ крайне динамична, и in vivo постоянно подвергается переформированию (с полупериодом <10 с!). В этом отношении белок напоминает тубулин эукариот.
У прокариот деление происходит либо путем образования перетяжки, либо с помощью перегородки.Для простоты, капсульный слой клеточной оболочки не показан.
В Z-кольце с белком FtsZ непосредственно взаимодействует белок FtsA, функция которого, вероятно, состоит в стабилизации кольца. Белок FtsA напоминает актин клеток эукариот, однако обладает дополнительным доменом, функции которого неизвестны. Этот белок образует димеры, но, по-видимому, не полимеризуется. Хотя он не участвует в формировании Z-кольца, клетки двойного мутанта, дефектного по белкам FtsA и ZipA, не способны образовывать кольцевые структуры. Таким образом, функции белков FtsA и ZipA частично перекрываются, и, по крайней мере, один из них необходим для стабилизации Z-кольца. Также показано, что белок ZipA непосредственно взаимодействует с FtsZ и, в отличие от последнего и FtsA, представляет собой трансмембранный белок. Поэтому ZipA может обеспечивать сопряжение Z-кольца с клеточной мембраной.
Остальные белки деления представляют собой трансмембранные белки. Функции белков FtsL и FtsQ неизвестны. Белок FtsW, вероятно, поставляет предшественники для белка FtsI, который является ферментом, участвующим в синтезе перегородки. Последний обладает способностью связывать пенициллин и взаимодействует с аппаратом синтеза клеточной стенки, функционирующим при делении. Белки FtsK и FtsN необходимы для деления клеток Е. coli, однако у B. subtilis гомолог белка FtsK (SpoIIIE) не участвует в делении, а гомолог белка FtsN у этих клеток отсутствует.
Между двумя хорошо изученными микроорганизмами , Е. coli и В. subtilis, существуют интересные различия в процессе сборки белков деления. Так, у E. coli этот процесс носит почти линейный характер, в то время как у В. subtilis сборка белков на Z-кольцевой структуре является взаимозависимой. Эти различия, вероятно, отражают различную организацию клеточной оболочки у грамотрицательных и грамположительных микроорганизмов. Пока мы мало знаем о том, каким образом полностью собранный аппарат деления влияет на цитокинез, и выяснение этих вопросов представляет собой обширное поле деятельности для исследователей.
Деление контролируется, главным образом, на уровне образования кольца FtsZ. Предполагают, что положение сайта деления, и, вероятно, протекание этого процесса во времени находятся под контролем двух факторов: блокирования нуклеоидом и системы Min. Оба этих фактора обеспечивают наступление деления только после завершения репликации ДНК, а также одинаковую величину образующихся клеток.
Фактор блокирования нуклеоидом исследован недостаточно. Он проявляется в том, что из-за своего объема нуклеоид может предотвращать деление. Поэтому деление клетки происходит только после завершения раунда репликации ДНК и расхождения сестринских хромосом с образованием отдельных нуклеоидов. При блокировании процессов репликации или сегрегации, присутствие нуклеоида в середине клетки предотвращает образование перегородки. В принципе отрицательный эффект нуклеоида может объясняться просто отсутствием в этой области исключением из его состава белка FtsZ. При этом белок не накапливается до критической концентрации, необходимой для его полимеризации.
Значимость фактора блокирования нуклеоидом для клетки представляет собой потенциальную проблему, которая заключается в том, что полюса клетки (по крайней мере у палочковидных бактерий) не защищены нуклеоидом, и поэтому возможно наступление аберрантного полярного деления. Для предупреждения этого, у многих бактерий присутствуют белки, входящие в систему Min, которая препятствует делению на полюсах.Название этой системы происходит от названия мини-клеток, образуемых мини-мутантами, для которых характерно деление на полюсах.
Ключевой эффектор системы Min представляет собой ингибитор клеточного деления, который называется MinC. Этот белок обладает способностью ингибировать образование Z-кольца, вероятно, непосредственно ингибируя полимеризацию FtsZ. Активность MinC находится под контролем белка MinD. Вероятно, этот белок контролирует внутриклеточную локализацию MinC по двум различным механизмам. Один из них состоит в том, что MinD транспортирует MinC на периферию клетки (ближе к цитоплазматической мембране) туда, где происходит сборка кольцевой структуры FtsZ. Второй механизм заключается в том, что MinD ограничивает активность MinC полюсами клетки, тем самым предотвращая наступление полярного деления, но способствуя делению клетки по средней линии.
У многих палочковидных бактерий система MiniCD используется для контроля за местонахождением сайта деления. Эта система хорошо охарактеризована у бактерий Е. coli и В. subtilis. Интересно, что у двух этих микроорганизмов существуют совершенно разные механизмы, посредством которых MinD ограничивает эффект MinC на полюса клетки. У В. subtilis используется простой механизм, при котором полярный якорный белок DivIVA транспортирует комплекс MinCD к полюсам клетки и в течение всего клеточного цикла удерживает его там в статичном положении. Как показано на рисунке ниже, DivIVA и MinD локализуются у полюсов вновь образованной клетки, и присутствие ингибитора MiniC предотвращает формирование FtsZ-кольца у полюсов.
По-видимому, после завершения репликации ДНК , в середине клетки создается новый потенциальный сайт деления. Концентрация ингибитора MiniC у полюсов позволяет провести сборку FtsZ-кольца в середине клетки и обеспечивает мобилизацию других белков деления. В этот момент аппарат деления, вероятно, становится нечувствительным к ингибирующему действию MinC Затем белки DivIVA и MinD перемещаются на середину клетки. Поэтому, когда при делении образуется новая пара клеточных полюсов, DivIVA встраивается в новые полюса и образует новую область проявления ингибирующего эффекта MinCD. Когда произошло сокращение оболочки, наступает разборка FtsZ-кольца, однако DivIVA и MinCD остаются на вновь образованных полюсах, тем самым предотвращая деление на этих полярных сайтах.
Таким образом, транспортировка DivIVA к сайту деления и затем его удержание на полюсах клетки являются ключевыми событиями этого механизма.
Интересно, что белок DivIVA локализуется на сайтах деления, когда он экспрессируется в эукариотических клетках (делящиеся дрожжи). Эта позволяет предполагать, что DivIVA может узнавать топологические характеристики, например кривизну мембраны, а не специфические белковые мишени.
В противоположность этому, в клетках Е. coli существует динамическая система MinCD, которая на какое-то время собирает комплекс у одного полюса. Затем он разбирается и собирается вновь у противоположного полюса. Так повторяется много раз. Этим процессом управляет кольцо белка MinE, которое, в свою очередь, каждый раз перемещается к тому или иному полюсу, смещая MinCD и обеспечивая ему возможность собраться у противоположного полюса. Изменение локализации MinCD от одного полюса к другому происходит с частотой порядка десятков секунд. Как показано на рисунке ниже, MinD поочередно накапливается на периферии мембраны с каждой стороны кольца MinE. Быстрое изменение локализации MinD не позволяет кольцу FtsZ собраться на полюсах.
Присутствие MinE в центральной области исключает проявление там ингибирующего эффекта MinD и дает возможность собраться в этом месте кольцу FtsZ. Остается невыясненным, почему для контроля MinCD и установления полюсов у Е. coli выработался такой энергетически невыгодный механизм.
MinD относится к интересной группе белков, обладающих общей функцией связывания нуклеотидов, которая также включает белок разделения хромосом, ParA. Близкий к ParA белок, Soj, также проявляет динамические свойства. Вероятно, общей для этих белков является их способность связывать и гидролизовать нуклеотиды и контролировать реакции полимеризации и деполимеризации. Это напоминает механизм контроля динамической нестабильности актиновых филаментов и микротрубочек у эукариот. Поэтому эти белки относятся еще к одному классу белков цитоскелета бактерий, обладающих широкими функциями, которые особенно связаны с вопросами морфогенеза на разных стадях клеточного цикла.
Недавно у грамположительных бактерий был идентифицирован белок, участвующий в блокировании клеточного деления нуклеоидом. Это Noc, представляющий собой белок, неспецифически связывающийся с ДНК, который локализован в нуклеоиде. Он также является ингибитором клеточного деления. Если не нарушена репликация хромосом, то мутанты noc растут нормальным образом. При этом в noc- клетках деление происходит с участием нуклеоида, а клетки дикого типа не делятся. Как показано на рисунке ниже, Noc и система MiniCD определяют местоположение кольца FtsZ в середине клетки. В клетках дикого типа, DivIVA запускает процесс полимеризации белка MinD, который распространяется от полюсов к середине клетки вдоль мембраны.
Белок MinC , связанный с белком MinD , предотвращает накопление FtsZ или полимеризацию поблизости от полюсов клетки. Предполагается, что белок Noc связывается с нуклеоидом и ингибирует накопление FtsZ или проявление его активности поблизости от нуклеоида. В клетках noc-, система Min предотвращает сборку кольца FtsZ, исключая область середины клетки, и клетки растут нормально. Однако у min- клеток Noc ингибирует сборку FtsZ только вокруг нуклеоида, и FtsZ образует кольцевую структуру в середине клетки и на полюсах, где нет нуклеоида. У клеток с отсутствующими топологическими ингибиторами (двойные мутанты min-noc-) сборке FtsZ ничего не препятствует, и по всей клетке образуются многочисленные вкрапления, состоящие из этого белка. Их образование приводит к утрате клеткой способности к делению. У грамотрицательных бактерий Noc отсутствует, однако у Е. coli обнаружен белок, контролирующий систему блокирования деления нуклеоидом по механизму, аналогичному Noc.